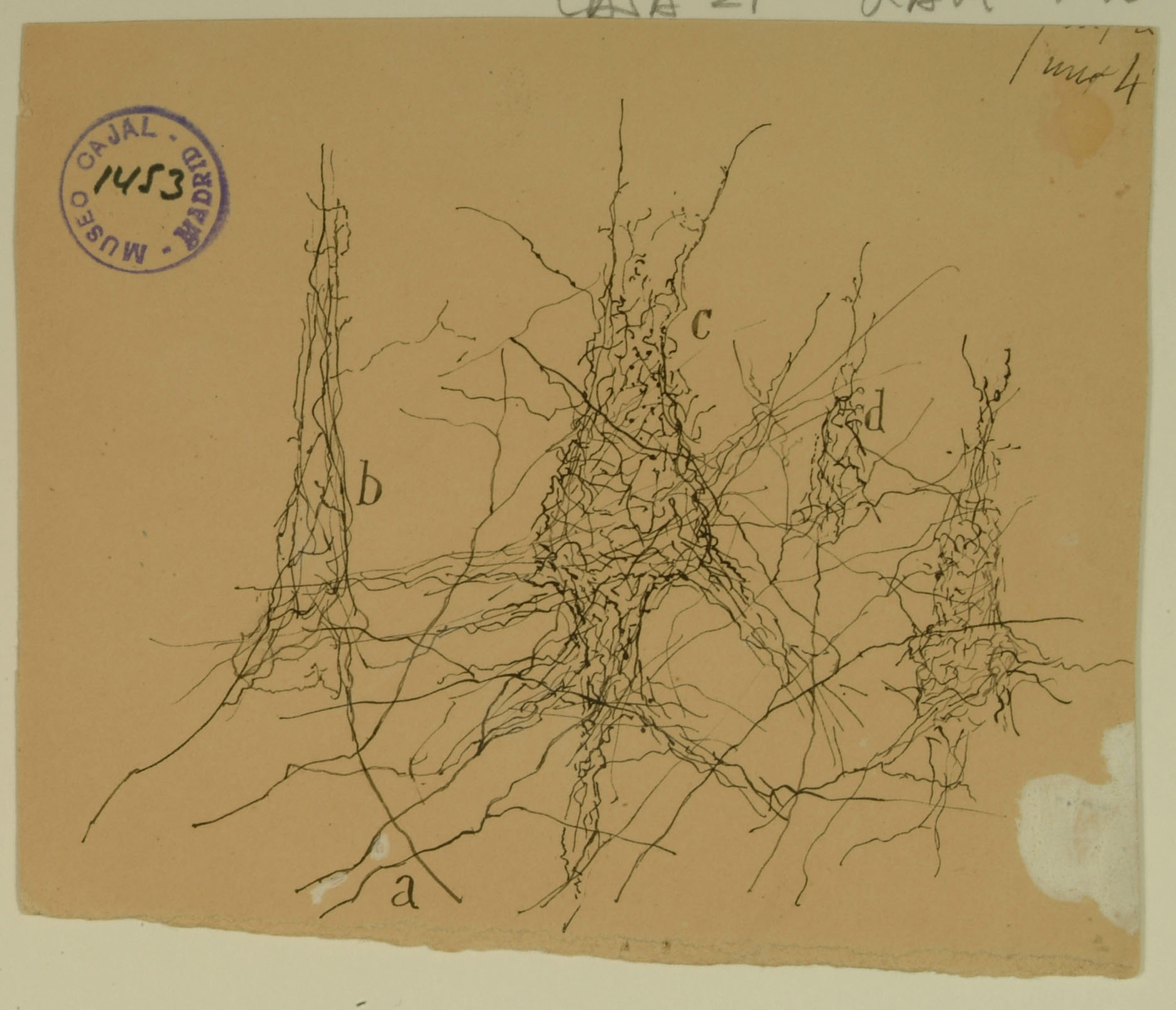
Formerly known as the ascending frontal gyrus, the precentral gyrus is the home of the primary motor cortex, which works in association with other motor areas to plan and execute movement. The primary motor cortex contains a variety of pyramidal neurons whose axons extend down the spinal cord and synapse with motor neurons. Shown above are pericellular nests around pyramidal cell bodies (b,c,d) in the primary motor cortex, formed by the profusely branched axons of other neurons. Cajal noted the presence of pericellular nests in the cerebellum, hippocampus, and cortex and hypothesized that they amplify signals from the presynaptic cells to the enveloped cell body. We now know that pericellular nests allow tight control of the targeted cell, crowding out input from other, more distant axons. While the precise function of the pericellular nests above remains unknown, recent research in the hippocampus has shown that they can influence memory and reinforcement of learned behaviors.
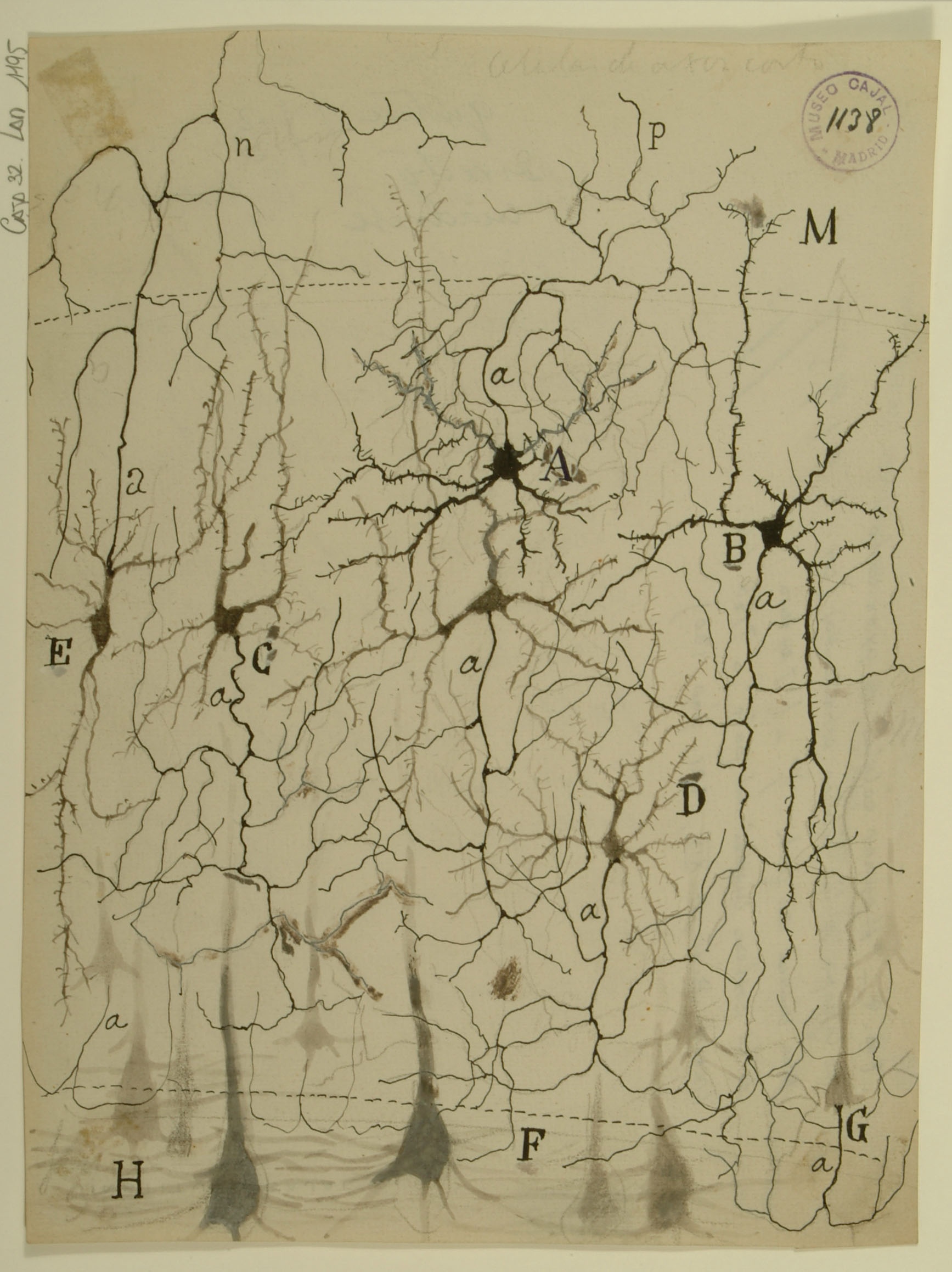
The primary visual cortex is the first processing center for visual signals within the brain, which is divided into six functionally distinct layers. Shown here are feline stellate cells (A,B) from Layer 4 of the primary visual cortex. Stellate cells receive the majority of their input from the lateral geniculate nucleus (LGN) of the thalamus, then send signals to pyramidal cells in layer 6 (H,F) for further processing. While studies of visual system gross anatomy dating from the 1940s indicated that information travels from the eye through the LGN and then into the cortex, more recent work has revealed that only 5-10% of the excitatory synapses onto stellate cells actually come from the LGN; most input comes from deeper cortical layers. The mechanism by which so few LGN inputs control stellate cells remains unknown – synapses from the LGN not appreciably larger than the others, nor are they clustered on stellate cell dendrites. One possible explanation is that signals from the LGN are highly synchronized with each other, whereas other inputs are more temporally dispersed.
The ventral cochlear nucleus is the first processing center for auditory signals in the central nervous system. Here, Cajal shows how individual auditory nerve fibers enter the cochlear nucleus ventrally as the cochlear nerve (A), bifurcating to form ascending and descending branches. The ascending branches (B) give rise to complex axonal terminals, the Calyces of Held, which synapse onto bushy cells to facilitate rapid, high-fidelity synaptic signals crucial to precise transmission of acoustic information. The descending branches travel posteriorly, giving rise to synapses in both the ventral (C) and dorsal (D) cochlear nucleus subdivisions that will perform more complex auditory processing, integrating feedback signals from deeper parts of the auditory pathway together with input from the cochlear nerve.
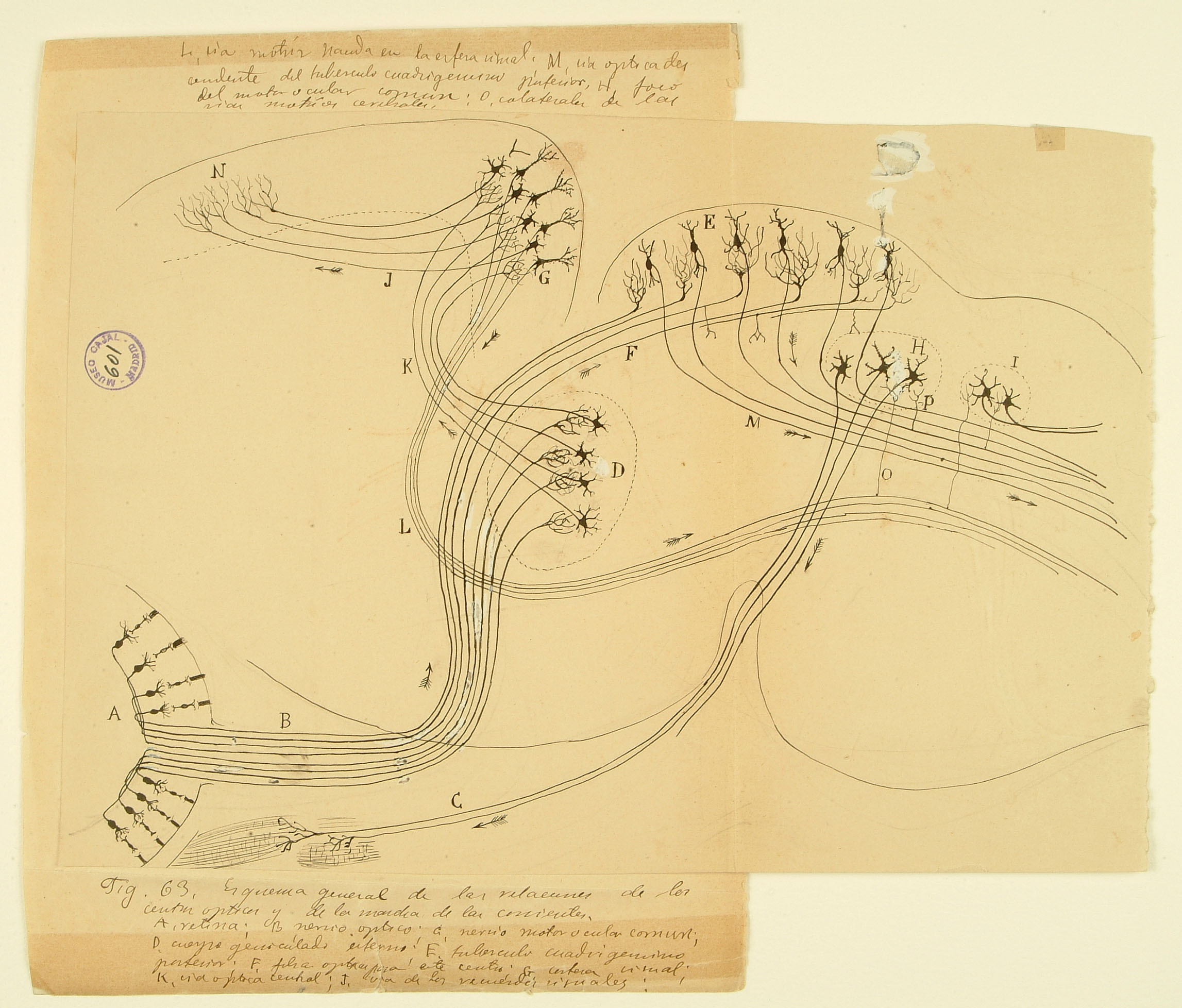
This drawing of the rodent early visual system illustrates one of Cajal’s most important contributions to science: the law of dynamic polarization, wherein information flows from the receptive dendrites of a neuron, through its cell body, and down through its axon, which then makes synaptic contacts onto dendrites of subsequent neurons in the pathway. Here he shows two routes by which visual information is used to control movements of the body. Signals from the retina (A) enter the brain by the optic nerve (B) and terminate in two different brain regions – the lateral geniculate nucleus of the thalamus in the forebrain (D) and the superior colliculus on the roof of the midbrain (E). The superior colliculus, evolutionarily conserved across mammal, integrates visual and other inputs to provide signals (M) instructing physical movement and orientientation. Neurons in the visual thalamus (D) relay signals to the visual cortex (G), which sends projections to higher-order cortical areas (N) and also provides another set of descending motor commands (L). This diagram was submitted by Cajal as part of a joint application for the 1902 Martinez y Molina Award titled “Sensory Centers in Man and Animals” with his brother Pedro, also a noted histologist who specialized in amphibian and reptilian brain structures.
Shown above is a schematic of the cochlea’s organ of Corti, named after the Italian anatomist Alfonse Corti, who originally described it in 1851. Its function is to transform the mechanical energy of sound waves into the electrochemical signals used by nerves via mechanosensory hair cells (G,C). Systematic variations in stiffness and size of the stereocilia arrayed atop each hair cell enable these specialized epithelial cells to respond to a range of sound frequencies. The signal is then transmitted to spinal ganglion neurons within the cochlear nerve (N) for processing in the auditory cortex. Hair cells are a popular subject of current studies, due to research into the mechanisms of their extremely sensitive transduction of mechanical stimuli. In addition, hair cells express many genetic mutations that underlie congenital deafness. Hair cell death is the primary cause of age-related hearing loss, prompting active research efforts to preserve or replace them.
The retina was a favorite subject for Cajal due to its accessibility, its orderly organization in alternate layers of cell bodies and intercellular contacts, and the easy identification of the main direction of nervous signal flow. Above is his first published illustration of the mammalian retina. His faithful depiction here highlights several structures that are now known or hypothesized to contribute to the optical properties of the tissue, in addition to their roles in visual signaling. The Müller cell (right portion of diagram), with its vertically oriented structure spanning the full thickness of the retina, has been proposed to guide light towards cone photoreceptors, thereby ensuring efficient photon delivery. Cajal also noted that the nuclei of rod photoreceptors (f) differ from those of their cone counterparts (e): animals with good night vision, like the dog from which this image was drawn, have evolved distinct chromatin patterns in their rod nuclei to facilitate light passage. In addition, Cajal highlighted in these photoreceptors large mitochondrial aggregates (l and n), which have been recently shown to act as microlenses concentrating light towards the absorptive photopigment in the outer parts of the cell.
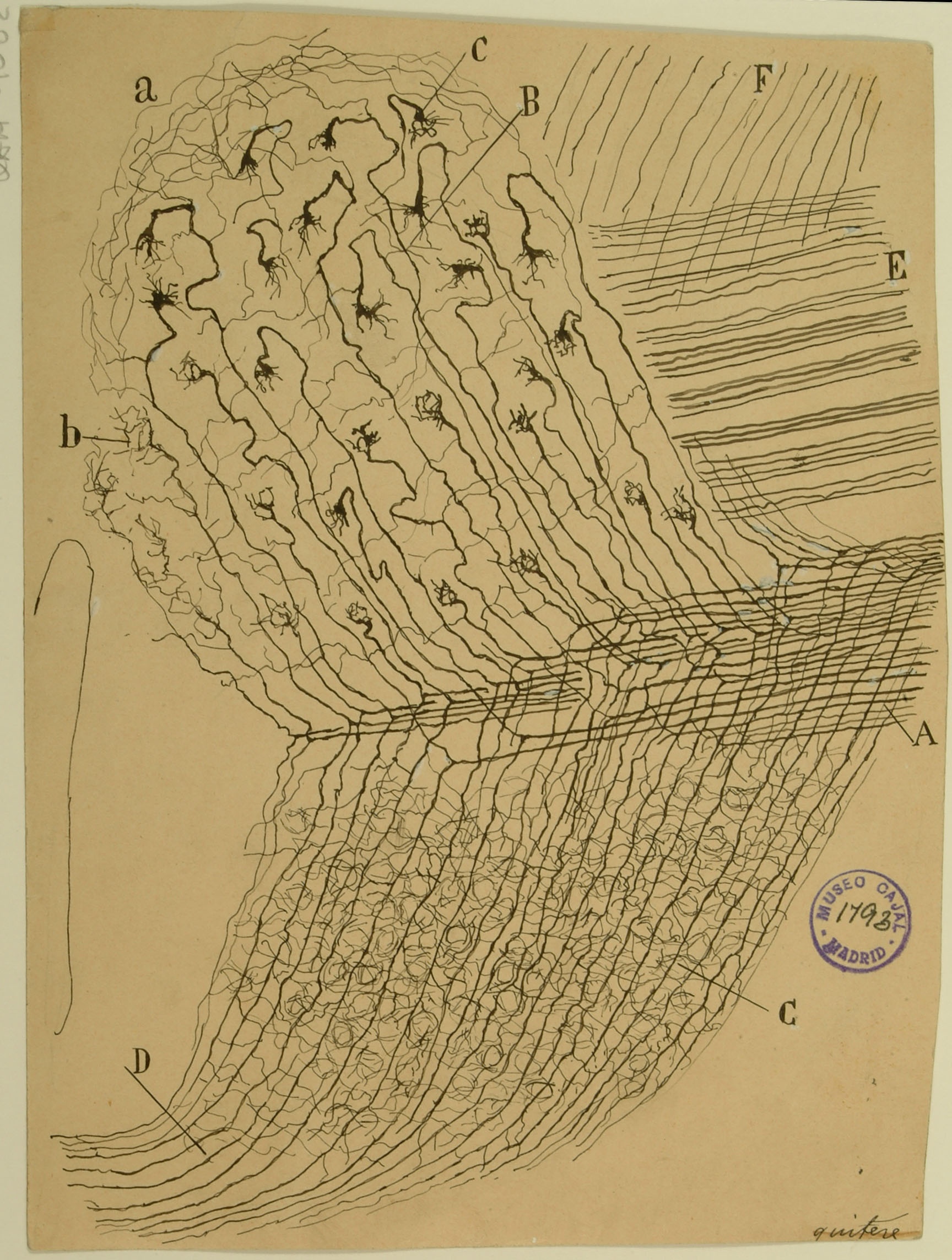
The cerebellum, an integral structure in transmitting sensorimotor signals in the brain, was the first system that Cajal studied. Through careful observation of its stained tissues, he first became convinced that neurons were distinct cells rather than part of a contiguous network. The cerebellum also served as one of the systems in which he would develop his law of dynamic polarization. The section shown here was taken from a sample perpendicular to the vividly branching Purkinje cell dendritic arbor that many think of when envisioning the cerebellum. The mossy fibers in the granular layer (C) transmit information from motor centers in the cerebral cortex to the granule cells (d), which send axons into the molecular layer (A). These axons split (e) to give rise to the parallel fibers (c), which contact Purkinje cells (b). Purkinje cells integrate these inputs in their large cell bodies, which occupy a distinct layer (B) and project axons to the deep cerebellar nuclei, where they provide inhibitory input. In more recent years, Cajal’s studies of the cerebellum have been corroborated by more modern techniques that have also refined our understanding of the fine organization of the cerebellum.